

Research
超巨大遺伝子群を制御するゲノム領域の発見
〜匂い受容体の遺伝子進化の謎の解明へ
(プレスリリース)
A Long-Range cis-Regulatory Element for Class I Odorant Receptor Genes
概要
東京工業大学 バイオ研究基盤支援総合センターの廣田順二准教授と岩田哲郎研究員、東京大学大学院 農学生命科学研究科の東原和成教授、理化学研究所 脳科学総合研究センター吉原良浩シニアチームリーダーらの研究グループは、匂いを感知する嗅覚受容体の遺伝子発現を制御する新たな転写調節領域(エンハンサー[用語1])を発見しました。
嗅覚受容体は、ゲノム上で最大の遺伝子ファミリー[用語2]を形成し、膨大な匂い情報の識別を可能にしています。嗅覚受容体ファミリーは、魚類から哺乳類に至る脊椎動物に共通して存在するタイプ(クラスI)と、陸棲動物特異的なタイプ(クラスII)に分類されます。そのうちクラスI遺伝子は、動物の進化の過程で染色体上の1カ所に留まり、長大な遺伝子クラスター[用語3]を形成しています。この長大なクラスI遺伝子クラスターの発現制御のメカニズムは長年、未解明でした。
今回本研究グループは、マウス遺伝学的手法と情報学的解析を駆使し、哺乳動物間で高度に保存されたクラスI遺伝子クラスターを制御するエンハンサーを同定しました。ゲノム編集技術[用語4]を用いた解析から、このエンハンサーは、この遺伝子クラスター全体を制御しており、制御する遺伝子の数とゲノム上の距離の両方において、これまでに例をみない規模で遺伝子発現を制御していることが明らかになりました。この”超”長距離作用性エンハンサー[用語5]の発見は、嗅覚受容体の遺伝子進化の謎を解く重要な成果と言えます。
この成果は、2017年10月12日(日本時間18:00)、英科学誌『Nature Communications』に掲載されました。
研究の背景
嗅覚は、外部環境中の匂い情報を感知・識別し、動物の生命維持のために必要な行動を起こす重要な感覚です。匂いを感知する嗅覚受容体は、ゲノム上で最大の遺伝子ファミリーを形成し、膨大な数の匂い情報の識別を可能にしています。その数はマウスで約1,100個(マウス全遺伝子の約5%で、ヒトでは約400個※ヒト全遺伝子の約2%)にも及びます。
嗅覚受容体には、「クラスI(水棲型)」と「クラスII(陸棲型)」の2つのタイプが存在します。クラスIは魚類から哺乳類まで脊椎動物に共通に存在するタイプで、クラスIIは陸棲動物特異的なタイプです。これまでの研究から、クラスIは水溶性の匂い分子を、クラスIIは揮発性分子を受容すると考えられています。クラスIとクラスII受容体遺伝子の染色体上の局在は大きく異なります。動物の進化の過程で、クラスI遺伝子が染色体上の1カ所に留まり続けて1つの巨大な遺伝子クラスターを形成しているのに対し、クラスII遺伝子は、遺伝子重複と転座を繰り返して、ほぼすべての染色体上に広がって分布しています。
遺伝子の発現には、個々の遺伝子の配列だけではなく、転写調節配列が必要です。これまでに、いくつかのクラスII遺伝子の転写調節配列が同定されてきましたが、ゲノム上最大級の遺伝子クラスターを形成するクラスI遺伝子の調節配列は、見つかっていませんでした。
研究の経緯
廣田准教授と岩田哲郎研究員らは、2013年にクラスI嗅覚受容体遺伝子の転写調節領域が特定のゲノム領域内に存在することを実験的に証明することに成功しました。今回この成果をさらに発展させ、マウス遺伝学的手法と情報学的解析を駆使することで、世界で初めてクラスI嗅覚受容体の転写調節領域の位置を突き止めました。
研究成果
(1)クラスI嗅覚受容体遺伝子クラスターを制御する超長距離作用性のエンハンサーの発見
重要な転写調節領域のDNA配列は、進化の過程で保存される傾向があります。そこで多様な動物種においてクラスI遺伝子クラスター領域の配列を比較し、保存されている配列を探索することで、新たな転写調節配列『Jエレメント』を推定しました。
生体内でのJエレメントの機能は、ゲノム編集技術を用いて作出した遺伝子改変(Jエレメントを欠失)マウスから明らかになりました。Jエレメント欠失マウスでは、75個(解析した遺伝子の58%)ものクラスI遺伝子の発現が減少し、その影響は遺伝子クラスター全体(約300万塩基対)に及んでいました。クラスIIのエンハンサーが約20万塩基対の範囲にある7〜10個の遺伝子を制御していることと比較すると、Jエレメントがこれまでにない規模で嗅覚受容体の遺伝子発現を制御していることがわかりました(図1)。また、これまでに同定されたどの遺伝子のエンハンサーと比べても、Jエレメントは制御する遺伝子数ならびに作用範囲が最大でした。Jエレメントの制御の範囲内に留まることでしかクラスI遺伝子は機能できず、このことが進化の過程でクラスI遺伝子が単一の遺伝子クラスターに留まり続けた要因になっていると考えられます。

図1 長距離作用性エンハンサーの作用距離の比較
βグロビン遺伝子のエンハンサーは、約6万塩基対の範囲内にある5つの遺伝子の発現を制御している。クラスII嗅覚受容体遺伝子エンハンサーは、約20万塩基対の範囲内にある7〜10個の遺伝子の発現を制御している。Jエレメントは、約300万塩基対内にある75個の遺伝子のエンハンサーと機能しており、長大な作用範囲を有している。
(2)嗅覚受容体の遺伝子発現メカニズムに新たな知見
1つの嗅神経細胞は、膨大な数の嗅覚受容体レパートリーから1つの遺伝子のみを選択し、発現します。これまで嗅覚受容体遺伝子の発現メカニズムとして、確率的に1つの遺伝子が直接選択されるモデルが提唱されています。しかし今回、Jエレメント欠失マウスの解析から、もう1つ新たな知見が得られました。従来のモデルが正しいとすると、対立遺伝子の片方のJエレメントを欠失したヘテロ欠失マウスでは、転写を活性化するエンハンサーが2つから1つになったため、クラスI遺伝子の発現数は半減すると予想されます。しかしながら、結果は予想に反して、クラスI遺伝子の発現数は半減せず、むしろ正常なマウスに近い発現を示しました。これは、片側のエンハンサーを欠失しても、もう一方の正常なエンハンサーが遺伝子発現を補填したことによります。このことは、遺伝子本体の選択より先に機能的なエンハンサーの選択が先行することに他なりません。以上のことから研究グループは、嗅覚受容体遺伝子発現のメカニズムとして、単一の遺伝子選択の前にエンハンサーが選択され、続いて選択されたエンハンサーの制御下にある嗅覚受容体遺伝子が1つ選択されるという新たなモデル(図2)を提唱しました。
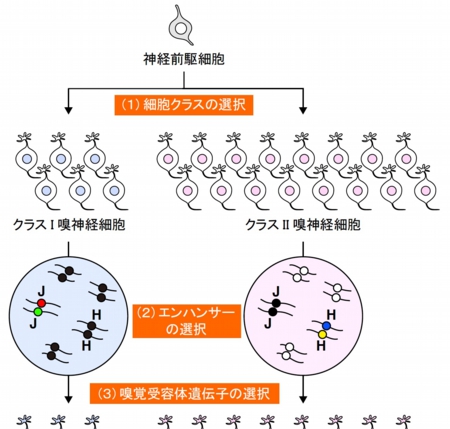
図2 嗅神経細胞における嗅覚受容体遺伝子選択モデル
(1)神経前駆細胞からクラスIもしくはクラスII嗅覚受容体遺伝子を発現する2種類の細胞が産生される。
(2)クラスIタイプでは主にJエレメント(緑と赤)が、クラスIIタイプでは多数のクラスIIエンハンサー配列から1つ(例:Hエレメント(青と黄)が選択される。
(3)選択されたエンハンサーの制御下にある1つの嗅覚受容体遺伝子が選択され、発現する。対立遺伝子座の片方のエンハンサーが欠失すると、決められた細胞集団を埋めるため他のエンハンサーが選択される(点線下段、J欠失(ΔJ)もしくはH欠失(ΔH))。クラスIタイプでは対立遺伝子座のJが優先して選択されるが、クラスIIタイプでは残りからランダムに選択されるためHを選択するものは見かけ上約半数となる。
今後の展望
クラスI嗅覚受容体のエンハンサー、Jエレメントの発見によって、長年未解明だったクラスI遺伝子の発現制御メカニズムの研究がさらに進むものと考えられます。また、陸棲動物特異的なクラスII嗅覚受容体と比較して、進化の過程で魚類から哺乳動物に共通に存在するクラスI嗅覚受容体の生理機能はほとんど明らかになっていません。今後、クラスI嗅覚受容体遺伝子の発現が大きく減少するJエレメント欠失マウスの匂い行動や生理学的解析によって、その機能が明らかになっていくと期待されます。
用語説明
[用語1]エンハンサー:ゲノム上には、タンパク質をコードする遺伝子領域のほかにも様々な機能を持つ配列が存在する。その一つである「エンハンサー」は、遺伝子発現を促進する配列として、遺伝子が発現するタイミングや組織を規定する。
[用語2]遺伝子ファミリー:進化上同一の祖先遺伝子に由来すると考えられる、配列や機能が類似した遺伝子群。嗅覚受容体遺伝子ファミリーは、7回膜貫通型Gタンパク質共役型受容体という特徴を共有する巨大ファミリーである。
[用語3]遺伝子クラスター:同様の機能を有する多数の遺伝子が、染色体上の同じ位置に直列して位置している状態(遺伝子集団)。クラスI嗅覚受容体遺伝子クラスターは、129個の遺伝子が約300万塩基対の範囲内に密集して存在する、極めて巨大な遺伝子クラスターの1つ。
[用語4]ゲノム編集技術:DNA分解酵素にゲノム上の狙った場所の配列を認識させることでDNAを切断し、目的遺伝子の配列を破壊したり、書き換えたりする技術。
[用語5]長距離エンハンサー:遺伝子から遥かに遠方に存在するエンハンサー配列。これまでに数十万〜百万塩基対離れた遺伝子に作用するものが報告されている。